
Con la entrada anterior de esta serie sobre la Biografía de la Vida habíamos cerrado el paseo a lo largo de una extenso periodo de tiempo de 4.038 millones de años, que agrupa a los tres primeros eones de nuestro planeta. A partir de hoy ya sólo vamos a movernos por el último de ellos, el Fanerozoico, el cual iremos despiezando con más meticulosidad dada la cantidad de información que disponemos de este periodo. No en balde es la etapa más próxima y por tanto con más datos disponibles.
El Fanerozoico se divide en tres grandes eras, Paleozoico, Mesozoico y Cenozoico, siguiendo el orden geológico de más antigua a más moderna. Cada una de ellas, a su vez, se divide en periodos. Hoy comenzaremos con el primero del Paleozoico: el Cámbrico, que se prolongó desde hace 542 hasta hace 488 millones de años.

Los primeros restos geológicos de esta época se estudiaron en Gales, por eso le nombraron Cámbrico pues en gaélico a Gales se le llama Cambria. El continente Panotia se estaba descomponiendo y la rica fauna Ediacara había desaparecido dando paso a la “explosión” de unas nuevas formas de Vida con características que se han prorrogado hasta la época actual. Los organismos son exclusivos del mar: bacterias, hongos, plantas y animales. Estos últimos van a descubrir el uso de materiales sólidos para crear protección y aportar sostén a sus blandos organismos.

La ruptura de Panotia y posterior deriva continental hizo que en esta época hubiera un gran continente, Gondwana, en posición ecuador-polo sur, del que se habían desgajado por el oeste los continentes de Laurentia, Siberia y Báltica, en posiciones ecuatoriales y tropicales, que llevaban su propia deriva de separación motorizada por una dorsal oceánica en forma de Y invertida. Toda la costa norte de Gondwana era una zona de subducción en donde iban creciendo arcos de islas volcánicas.
No se sabe muy bien cuál pudo ser el clima del Cámbrico aunque se supone que tuvo temperaturas cálidas, ni muy altas ni muy bajas. Los abundantes sedimentos calizos parecen asegurar esta situación climática. En todo el periodo no hay evidencias de glaciaciones, lo cual refuerza la hipótesis de temperaturas benignas.
La Vida continuaba en los mares. Sus fondos más profundos estarían recubiertos de sedimentos silíceos y calcáreos. En la proximidad de la línea costera estaba el roquedal, firme o disgregado. Los arrecifes hacía tiempo que habían aparecido en la franja de aguas tropicales en donde habían adoptado formas globulares, de abanico o tubulares, más o menos delicadas según la profundidad. Al comienzo de esta época estaban formados por esponjas o animales parecidos, arqueociatos y algas calcáreas. Más tarde a estos tres tipos de organismos se les unirían los corales. Se seguían construyendo formaciones de estromatolitos, aunque ahora no con tanta facilidad debido a que alguno de los nuevos animales que iban apareciendo ya comenzaba a alimentarse de sus capas bacterianas. La fauna Ediacara había desaparecido hacía unos millones de años y no se disponen de muchos restos fósiles que la enlacen con la cámbrica. Quizás podamos considerar a los individuos de los géneros Parvacorina y Springgina como los que firmaron la transición, y que suponemos pudieron ser los ancestros de los trilobites, unos artrópodos de la gran explosión de vida a la que estaba llegando. Sabemos también que las esponjas estaban allí, pero no mucho más. A lo mejor la vida estaba constituida por seres de cuerpo blando, con una probabilidad muy baja de fosilizar, o eran microscópicos. O quizás aún nos falta por descubrir el yacimiento fósil definitivo que complete la línea evolutiva.

Spriggina flounensis, animal de finales del Ediacárico que se postula como enlace con los artrópodos del Cámbrico (Wikimedia, CC BY 2.5)
Los fondos marinos y las orillas someras debían estar cubiertos de tapetes bacterianos y de algas. Se acepta de manera general que no existían plantas sobre la tierra en esta época. Aunque la datación molecular nos sugiere que las plantas terrestres más simples se segregaron pronto del árbol evolutivo, en el Precámbrico, hace unos 700 millones de años, y los hongos hace 1.000 millones de años, también en el Precámbrico. Es posible que un manto formado por hongos, algas y líquenes ya cubriera ciertas zonas de la Tierra. Muchas incógnitas y mucho por descubrir de la Vida en estos momentos.
En poco tiempo el panorama cambió.
La expansión de la biosfera
A principios del Cámbrico se produce una tremenda y rápida diversificación de formas vivas. Un fenómeno que hoy se sabe fue de carácter global en todo el planeta y que se produjo a lo largo de un periodo geológicamente mínimo: unos diez millones de años. Se la conoce como la explosión del Cámbrico.
A través de un juego de éxitos y errores, que iban siendo permanentemente calibrados por criterios evolutivos, se inventaron nuevas estrategias de alimentación, pasando de pacer microorganismos al azar a una búsqueda activa del alimento. Los niveles de oxígeno en las aguas habían sobrepasado el límite necesario para que los organismos pudieran metabolizar colágeno, proteína fundamental para disponer de un organismo rígido y flexible: la fauna ediacara podía ser superada. También con esos niveles de oxígeno los organismos podían aprovechar el calcio y el silicio disueltos en el medio para usos particulares casi inéditos. Los organismos se van a ir haciendo más complejos que las sencillas esponjas. Recordemos sus flagelados coanocitos. La naturaleza aprovechó la oportunidad de fortalecer los simples cilios, incluso uniéndolos formando una membrana que sería vibrátil, con lo que se inventaba un sólido aparato locomotor: la carrera armamentística para la predación acababa de hacer acto de presencia.
El buen clima y la fuerte disgregación de terrenos continentales estaban favoreciendo la precipitación de calizas en los mares y el “envenenamiento” de sus aguas con iones perjudiciales para el metabolismo, lo que impulsó a los organismos vivos a desarrollar un bombeo de estos minerales fuera de sus cuerpos. Por este procedimiento a algunos se les fueron desarrollando en su superficie escamas y espículas calcáreas o silíceas que inicialmente les ayudarían a protegerse. Quizás como subproducto, y gracias a que en el proceso pudo generarse un diseño mecánico que acabó formando esqueletos tanto internos como externos, la acumulación de iones contribuyó a mejorar los elementos de sostén y a que la movilidad pudiera realizarse con mayor soltura. Al permanente espolvoreado calizo y silíceo de las aguas se le unió el aprovechamiento de biopolímeros tales como la quitina y el colágeno. Caparazones, cáscaras, espículas, tejidos más consistentes, cartílagos… aparecieron por doquier durante el Cámbrico. Fue el inicio de una gran carrera armamentística: a un predador más hábil le sucedía una presa más protegida, que obligaba a incrementar las habilidades de los predadores… y así en continua progresión. Patas, ojos, mandíbulas,… un sinfín de nuevas armas. Éste fue el gran motor de la expansión de vida en el Cámbrico: se pasó de esperar a que el alimento viniera gratuitamente a buscarlo activamente.
Los primeros escalones de la cadena alimentaria comenzarían en las algas y los tapetes bacterianos, entre los cuales pulularían pequeñas larvas de animales. Los suelos oceánicos estarían repletos de limos con detritos, restos de la vida que sobrenadaba. El plancton flotante o suspendido entre dos aguas había sido abundante con anterioridad. Pero en estos momentos estaban apareciendo nuevos animales con ojos para decidir dónde ir, con asomo de mandíbulas masticadoras que les permitían pacer en las praderas submarinas, animales con rudimentarias paletas con las que escarbaba en el limo, seres con facilidades para reptar o nadar que cubrían cualquier nicho en donde pudiera haber alimento. Gusanos que se enterraban en el fango, gusanos con numerosas cerdas móviles, artrópodos acorazados y con terribles “mandíbulas”, cefalópodos ágiles y voraces, todos conviviendo con los estáticos moluscos.
Según comenta el paleontólogo norteamericano Stephen Jay Gould acerca de la explosión del Cámbrico, parece como si en aquel momento la Vida manifestara libremente su potencial, haciendo realidad un inmenso abanico de posibilidades, todas con exquisita funcionalidad. Realmente fue como abrir una caja de sorpresas en donde se encontraban almacenadas una gran variedad de piezas dispuestas para ser utilizadas, para diseñar y montar cada una de las partes vitales de un organismo: cien formas de boca, cien de patas, cien de cabezas… mézclese y tómese una de cada y hágase un animal cámbrico. Las variantes montadas que sean viables encontrarán su encaje en el medio ambiente adoptando un hábitat particular y una posición en la cadena trófica. Desde el fondo marino a las aguas intermedias, desde ser detritívoros a ser auténticos depredadores. Una panoplia completa. Y todo en el “suspiro” de tan sólo diez millones de años.
Parece como si en aquel momento cualquier solución, en cuanto a formas y funciones, que tuviera posibilidad de éxito era inevitable y tenía que surgir espontáneamente en la naturaleza. Es el sino de la evolución ya que sin duda el esplendor de la variación siempre se repetirá, con mayor o menor ímpetu, tantas veces como lo ordenen las circunstancias de cada momento. Algo parecido a lo que estamos viendo durante el Cámbrico sucedió unos 500 millones de años más tarde en el Cenozoico, cuando África, Sudamérica y Australia navegaban a la deriva como inmensas islas solitarias. Sobre sus geografías las faunas de mamíferos, en unos casos placentarios y en otros marsupiales, evolucionaron de forma independiente y ¿sorprendentemente? casi siempre adoptando soluciones convergentes. En cada uno de estos continentes la evolución llevó a colonizar nichos semejantes con animales de fenotipos semejantes y ello sin tener ninguna conexión familiar inmediata. El lobo euroasiático y el lobo de Tasmania, el topo y su homólogo marsupial, el oso hormiguero y el numbat marsupial australiano, el armadillo sudamericano y el pangolín afroasiático, y así muchos más ejemplos. La Vida es una activa maquinaria llena de iniciativa que en donde encuentra un resquicio, allí se instala.
Es muy posible que todo se originara con los Pequeños Fósiles con Concha (Small Shelly Fossils) conocidos también como la fauna Tomotiense, de hace 543 a 530 millones de años. Prácticamente son los primeros restos de esqueletos mineralizados que aparecen en el registro fósil, al comienzo del período Cámbrico. Los restos consisten en escamitas, espículas, tubos y conchas, que bien pudieran ser elementos de organismos mayores en una época de esqueletización temprana, aunque otros parecen ser conchas individuales pertenecientes a organismos de pequeño tamaño. Seguramente las rarezas estructurales de esta fauna corresponden a planes corporales que aparecieron rápidamente y murieron igual de rápido sin dejar supervivientes, como ya les había sucedido a los organismos de Ediacara. De hecho, vimos en la entrada anterior número 15 que, al igual que estos últimos, se extinguieron coincidiendo con una excursión negativa del δ13C.
No obstante no hay que menospreciar a las “pequeñas conchas” ya que en algunas de ellas ya se observan precursores diseños bilaterales. Incluso, un poco anteriores a ellos (550 millones de años), ya sabemos que había animales que se desplazaban según el eje mayor de su bilateralidad. Se trata del gusano, o lo que sea, Yilingia spiciformis, encontrado en el yacimiento chino de Dengying, en la región de las Tres Gargantas del río Yangtze.[1]

Variedad de los Small Shelly Fossiles (de izquierda a derecha, a partir de Fossil.wiki, Paleos.com, ambos fair use y wikimedia y wikimedia, ambos CC BY-SA 3.0)
Más allá de esta pequeña fauna las formas más avanzadas de organismos vivos se conocen en gran medida gracias a unos pocos yacimientos extremadamente ricos en fósiles. Uno de los más importantes es la “catedral” de Burgess Shale, en la Columbia Británica, en Canadá, sobre las laderas de las Montañas Rocosas, que contiene fósiles con antigüedad comprendida en la franja de 540 a 530 millones de años atrás. En aquellos momentos sólo el noreste de Canadá asomaba la nariz por encima del agua en el continente de Laurentia. El resto era el mar. Burgess Shale es un verdadero cuerno de la abundancia de formas animales, gracias a la que sabemos mucho de la fauna del Cámbrico. Artrópodos, cordados, gusanos, esponjas y otros animales desaparecidos.

Fauna de filos encontrados en Burgess Shale que en la actualidad no tienen representantes vivos: 1-Opabinia, 2-Viwaxia y 3-Yohoia (Wikimedia, wikimedia y wikimedia, GNU FDL 1.2)
Lo mismo podemos decir del yacimiento formado por los esquistos de Maotianshan en China. Su antigüedad es de 522 millones de años y en aquel momento la región estaba sumergida en la línea del ecuador. El inicio de su investigación es posterior a la de Burgess Shale y por tanto aún quedan muchos misterios por descubrir, aunque, como en el yacimiento canadiense, la diversidad es asimismo fantástica.
Los múltiples estudios realizados a partir de estos yacimientos han dejado encima de la mesa de la evolución una gran pregunta: ¿Cómo en tan poco tiempo, geológicamente hablando, aparecen todos los planes de diseño corporal de los organismos, entre ellos los mismos, y no más, de los que vemos en los animales de hoy en día? ¿Por qué en los siguientes 525 millones de años no se han innovado nuevos diseños y sólo se han modificado los que aparecen en los yacimientos del Cámbrico? ¿Por qué aparece el tipo de fósiles que aparece? ¿Cuáles fueron los eslabones contemporáneos de la fauna ediacara o de la de las pequeñas conchas, ambas extintas, que llegaron hasta las formas complejas del Cámbrico?
Hay teorías que explican esta rápida expansión de la diversidad apoyándose en el argumento de la presión de la predación y la competencia. El hecho de que hubiera cada vez más formas de animales complejos, que se iban armando defensiva y ofensivamente de forma progresiva, hizo que la biosfera animal buscara múltiples soluciones y salidas a esta presión, haciendo evolucionar rápidamente desde las formas simples los múltiples y diferentes planes estructurales, lo que llamamos filos, de los organismos de comienzos del Cámbrico.
Podríamos pensar también como idea general que a partir de un espacio vacío es más fácil el rellenarlo. No obstante algunos expertos se resisten a pensar, como es el caso de S. J. Gould, que el éxito de la “explosión” fuera tan sólo consecuencia del estado “virginal” y sin contaminación histórica en que se encontraba el medioambiente cuando todo se inició. Esto sólo sería una parte. En entradas posteriores de esta serie veremos que se dieron grandes extinciones, como la del Pérmico-Triásico en la que desaparecieron casi el 95% de las especies marinas, tras las que sabemos que la biosfera quedó casi despoblada, por lo que podemos pensar que de nuevo se habría generado un estado casi “virginal”. Y lo que es cierto es que después de este repetido barrido y casi puesta a cero no se observa ninguna nueva explosión de diversidad. Por tanto, no está tan claro que el extraordinario impulso que se produjo en el Cámbrico fuera sólo consecuencia de que los nichos ecológicos se quedaron vacíos, en una situación donde entonces pudo manifestarse la Vida con toda su potencialidad.
Debieron concurrir además otras circunstancias. Los genes de los pocos animales que iniciaron el camino tras el fracaso de los ediacarenses, debían mantener unas interrelaciones más sencillas que la que mantienen estos mismos tipos de genes hoy en día. Sus duplicaciones en el ADN con el paso de los millones de años y el posterior cambio funcional, enmarañó e hizo más dificultosas sus formas de expresión. Sin embargo, en aquella época debían ser más “libres”, estar menos condicionados, y por tanto con un alto potencial de expresar un variado escaparate de realidades. Por otro lado, el hecho de haber tenido que ser tan pocos los animales progenitores en las generaciones iniciales del Cámbrico nos permite pensar que el acervo genético de la época era “escaso” y compartido por todos los organismos que iban surgiendo de nuevo: todos dispondrían de parecidas estructuras de expresión fenotípica. De ahí la gran mezcla de soluciones particulares entre todos los nuevos modelos animales de inicios del eón Fanerozoico. Multitud de filos que compartían soluciones estructurales, a veces en distintas partes de sus organismos, dentro de unos patrones corporales que no tenían nada que ver unos con otro: patas articuladas, ojos articulados, cuerpos articulados, antenas articuladas.
Después de estos 10 millones de años iniciales ya no hubo más inventiva, sólo aniquilación de aquellas formas a las que en cada momento biogeológico las circunstancias les iban poniendo el cartel de “menos adaptadas”. Como ya hemos comentado, hoy en día hay muchos menos filos que los que se conocen a partir de los fósiles de la biota del Cámbrico. Menos pero también más singulares, en el sentido de que ahora prácticamente no se repiten fuera de un filo aspectos particulares de su fenotipo. Es como si la continua poda del árbol engendrado a principios del Cámbrico hubiera abierto tales huecos entre sus ramas que ya unas no se reconocen en las otras: gusanos, moluscos, artrópodos, braquiópodos, cordados… realmente son estructuralmente muy distintos.
El genetista John Maynard discurría al hilo de lo anterior que una vez establecido el escenario con tal riqueza de formas, no le quedó más alternativa que ir simplificándolas. En un medio con un catálogo de filos abarrotado, para que surgieran otros nuevos o bien tenían que aparecer a partir de mutaciones en los existentes, o bien tenían que surgir de nuevo, de la misma forma como lo habían hecho los ya existentes, es decir, a partir de células simples u organismos primitivos muy sencillos. En el primer caso, es decir, a través de mutaciones en los filos existentes,el salto de un filo a otro parece muy improbable dadas las diferencias tan grandes que hay entre los diseños de cada uno de ellos, de forma que la macromutación que hubiera llevado de uno a otro se hubiera hecho inviable. En la segunda hipótesis, la generación de nuevas estructuras desde escalones iniciales también parece improbable, ya que la competencia de los organismos más complejos, con una historia a sus espaldas y ya bien establecidos, anularían por selección natural a las nuevas aventuras. Así que a la biosfera animal sólo le quedó el camino del ajuste por regresión y disminución de variedad, como así ha sucedido.
La opinión de S. J. Gould se entrecruza con la de J. Maynard, a la vez que se apoya también en la ya comentada progresiva regresión de los grados de libertad de los genes. A medida que iban pasando los millones de años y la selección natural iba “cazando” a los que se despistaban, fueron quedando unos pocos cada vez más especialistas de sus nichos medioambientales, ya que en ello les iba la vida. Estos filos seleccionados, estos organismos con genes bien interrelacionados que les habían llevado al podio de la biota se encontraron con las manos atadas por su superespecialización, lo que les hacía muy difícil expresar grandes mutaciones. Así que el árbol fue proyectando sobre el horizonte unas ramas cada vez más potentes… y cada vez más escasas. Queda como la gran duda para Gould el determinar si las circunstancias que iban colocando a un determinado filo como último de su rama, al alcance de las tijeras podadoras de la selección natural, fueron en cada momento consecuencia de mil causas azarosas o bien resultado de un camino inevitable y predestinado. De los veinticinco filos fósiles catalogados en Burgess Shale sólo cuatro perduran entre nosotros, filos éstos que sin lugar a dudas no tenían por qué ser necesariamente los mejor preparados, más que los otros veintiuno. Fueron los triunfadores porque las cartas les vinieron de cara.
La historia evolutiva del Cámbrico
Aquellos mares de hace más de 500 millones de años no eran muy distintos a los de ahora. Poblados por las mismas líneas familiares de animales de hoy en día, pero con otros cuerpos. Nos hace pensar que aquellos animales, hoy fosilizados, también provenían de otros antecesores de los que no sabemos mucho. Entre las antiguas bacterias y ellos se extiende un casi vacío del que sólo podemos especular con los datos de los que disponemos: ya sean morfológicos, extrapolando e imaginando parecidos entre los escasos fósiles y sus tataranietos que hoy en día viven, o bien de tipo genético, mediante análisis moleculares de proteínas o del ADN de sus parientes actuales –o de fósiles, aunque esta molécula se degrada con facilidad- buscando semejanzas de genes y hasta cuánto se parecen con sus descendientes actuales y entre ellos. El hecho de que los primitivos protistas tuvieran el cuerpo blando de difícil fosilización no ayuda a seguir las pistas. Pero vamos a intentarlo.
¿Cuál pudo ser el guión de esta película? Rebobinemos un poco el celuloide y vayamos de nuevo a los últimos momentos del Precámbrico. Como ya hemos comentado, el árbol filogenético se abrió exageradamente durante aquel periodo. Por los registros fósiles vemos que casi al comenzar el Cámbrico las esponjas continuaban con una fuerza vital extraordinaria, mientras que las medusas y pólipos ya se buscaban la vida. No serían muy distintos de sus parientes actuales.
Hoy en día los dos últimos, medusas y pólipos, prácticamente comparten un mismo tipo de larva, llamada plánula, ciliada y nadadora, que tiene simetría bilateral. Puede optar por tener una vida libre flotando en el mar o bien anclarse en una roca del fondo. En la siguiente imagen podemos descubrir su anatomía no muy diferente a la de la imagen contigua, que sorprendente no es la de otra plánula sino la de un Acelomorfoactual. El paso ancestral de una a otra forma, de una larva de medusa a un animal bilateral, bien pudo ser debido a un proceso de “pedomorfismo”, palabra muy rara pero que sólo quiere decir que en algunos casos el momento de la madurez joven/adulto se va retrasando, pero no así la madurez sexual que se mantiene en el tiempo, de forma que en algún momento los animales reproductores maduros evolucionados llegan a fenotipos que mantienen la morfología juvenil de sus antepasados. Este pudo ser el mecanismo de enlace, unido también a un proceso de evolución convergente, entre los primitivos habitantes que sobrepasaron desde el Proterozoico la frontera del Cámbrico y los más básicos bilaterales sobre los que se gestó la fauna de la “explosión”, los antepasados de todos los animales actuales.

Plánula de vida libre (izquierda) (intrabiodev.obs) y Acelomorfo (derecha) (Fuente: Dr Bernhard Egger, UCL.)
Al actual Acelomorfo se le consideraba hasta hace poco un diminuto gusano platelminto, un pequeño ser que se encuentra en la base de los animales bilaterales. Actualmente se le ha asignado un filo propio separado del de los platelmintos por lo que toma más énfasis la frase anterior. Eso quiere decir que un tatarabuelo suyo posiblemente fuera el ancestro común de todos los bilaterales, la mayoría de los animales de hoy en día, y el primero de ellos que constituyó filo propio. Si acudimos de nuevo al árbol filogenético de Dawkins presentado ya hace dos entradas, la número 16, y hacemos un zoom sobre el momento del inicio de los animales bilaterales, nos encontramos con el siguiente esquema que nos permite asegurar la cercanía evolutiva de ambos filos:

Como podemos observar Richard Dawkins teoriza que el momento del inicio de la gran aventura, cuando se dio la separación entre cnidarios y acelomorfos, pudo haber sido hace ahora unos 700 millones de años. A partir de aquel momento un ser parecido a la plánula, ciliado y plano, con simetría bilateral, estaba predestinado a ser el patriarca de todo el pueblo animal que le sucedió. Y se tuvo que esperar unos 110 millones de años de remodelación desde este animal y su descendencia para llegar a un diseño, aunque primitivo, más acorde con nuestros patrones habituales: lo imaginamos como un gusano con simetría bilateral, primo de los platelmintos, con un extremo que hacía de cabeza, en el que se abría el orificio de entrada al estómago. Tendría también unos ojos rudimentarios, apéndices bucales que usaba para alimentarse, un cuerpo que apuntaba ya hacia una segmentación, aunque realmente no era así, y algún tipo de ornamentación corporal. “Dawkins dixit“.
Este tipo de humilde espécimen nos recuerda a la contemporánea Planaria y sería el cabeza de serie, el ancestro común, de la fauna del Cámbrico. Vamos a ir ahora por barrios, guiados por el siguiente plano callejero con el que hoy en día se encuentran más confortables todos los informes de genética molecular.
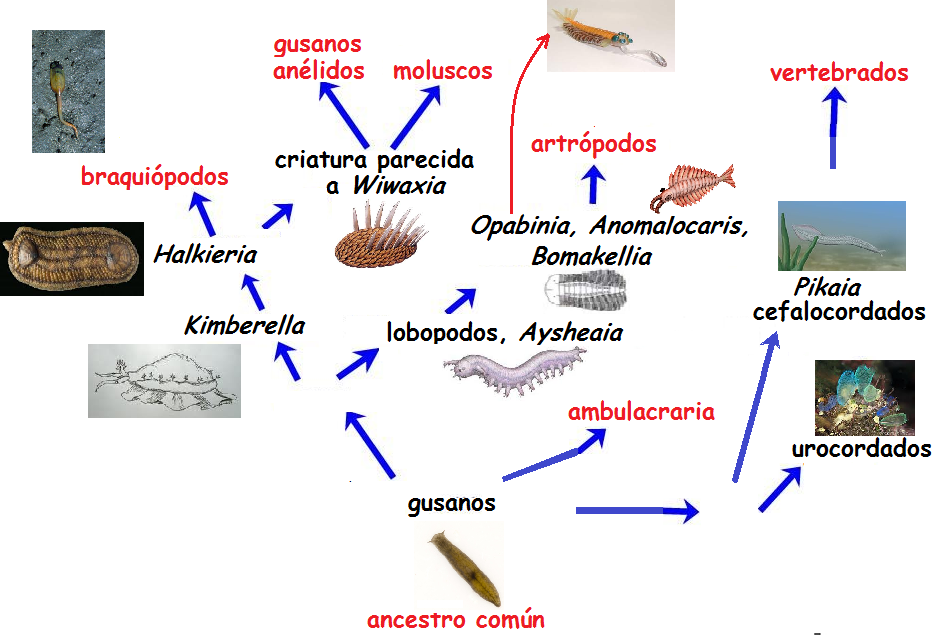
Caminos evolutivos de los especímenes de la fauna del Cámbrico. La imagen del ancestro común corresponde a una Planaria actual
Esto se pone interesante. La figura anterior es la hoja de ruta que vamos a seguir a lo largo de varias entradas a partir de ésta. En la próxima nos atreveremos a analizar el inicio de todos ellos. Nos esperan los artrópodos, braquiópodos, moluscos, anélidos y vertebrados… en definitiva, los principales filos de la biosfera. Nos vemos entonces.
- Más información en el siguiente artículo, o en este más para legos en la materia, presentado en la revista Nature de septiembre de 2019. [↩]

The La Biografía de la Vida 18. La biota del Cámbrico by , unless otherwise expressly stated, is licensed under a Creative Commons Attribution-Noncommercial-No Derivative Works 2.5 Spain License.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{ 3 } Comentarios
Jaime, la serie es fascinantey tu generosidad al compartir este enorme trabajo es una de esas conductas que evidencia que en la evolucion triunfa la cooperacion frente al egoismo. Un abrazo
Hombre Gabi, mira por donde me sale un muy buen viejo amigo y compañero de trabajo en este blog. Una alegría.
Me alegro que te guste y que disfrutes con la serie, lo mismo que disfruto yo con el tema desde hace unos años. Conocer es libertad mental. Si el tiempo acompaña, como decían los taurinos, hay serie para rato. Se aceptan críticas de todos los estilos pues para mi siempre suponen una “mejora”.
Y para enorme trabajo, el tuyo. Gracias a tu gestión se iban resolviendo cantidad de líos personales, siempre de forma eficiente y amable. Un fuerte abrazo.
El artículo me ha aclarado mucho sobre la evolución rápida en el Cámbrico. 10 millones de años en los períodos geológicos, es un tiempo breve
Escribe un comentario