
Siguiendo el recorrido fijado por esta serie sobre la Biografía de la Vida hoy nos toca avanzar hacia un nuevo periodo geológico. En la entrada anterior nos quedamos en el año 416 millones, anterior al día de hoy, por lo que hoy nos toca dar un paso más y adentrarnos en el Devónico, que duró hasta hace 359 millones de años. Su nombre procede de Devon, un condado ubicado en la península de Cornualles, en el suroeste de Inglaterra, donde las rocas de este periodo son muy comunes.

A este periodo le podemos poner la etiqueta de “el de las tres migraciones”: las plantas se extienden por la tierra llegando a formar enormes bosques; los artrópodos la colonizan fundando la estirpe de insectos y arácnidos; y los peces, después de su evolución en el agua, siguen sus firmes pasos hacia tierra firme.

La configuración de los continentes es aún semejante a la del Silúrico. Gondwana dominando la posición austral mientras Siberia migraba hacia el norte. Entre los dos se consumaba la unión de Laurentia, Báltica y Avalonia que formaron sobre el ecuador el provisional continente de Euramérica. Su posición geográfica le confirió una climatología seca, que se tradujo en unas zonas prácticamente desérticas en donde se formaron los antiguos depósitos sedimentarios de arenisca roja, compuesta por hierro oxidado de color rojo características de las zonas secas. Es por eso que a Euramérica se le conoce también como el continente de las Viejas Areniscas Rojas.
La orogenia Caledónica completa su estructura y es seguida, al ir cerrándose la proximidad entre Laurasia y Avalonia, por la orogenia Acadia que formó los montes Apalaches, actualmente en la costa este de Estados Unidos.
Poco a poco se iba acercando Gondwana, que empujaba desde su posición sur, hacia Euramérica, firmemente asentado en su posición ecuatorial, haciendo desaparecer progresivamente el océano Rhéico, al sur. Al mismo tiempo la deriva norte de Siberia le dirigía hacia posición de colisión con Euramérica, haciendo cada vez más angosto el océano Ural.
El nivel del mar era elevado en todo el mundo y gran parte de las tierras estaban sumergidas bajo aguas someras donde vivían los organismos de arrecifes tropicales. La intensa actividad de construcción de estos arrecifes indica que el clima devónico fue en general cálido y estable. Sin embargo, la proliferación de las plantas en tierra firme introdujo un nuevo vector en la fotosíntesis, incrementándose la absorción de CO2 atmosférico y por tanto disminuyendo la capacidad invernadero de la atmósfera.

Quizás por ello a finales del Devónico comenzó un progresivo enfriamiento de la Tierra. Como resultado del incremento de la fotosíntesis, en la atmósfera la concentración de oxígeno se incrementó, acercándose a los niveles actuales del 21%.
Las plantas
Durante este periodo el paisaje terrestre cambió drásticamente. La paleta de colores se orientó hacia el verde gracias al desarrollo de las plantas. Dejamos en el Silúrico algunas breves pinceladas próximas a las corrientes y charcas de agua, provocadas por los organismos que se esforzaban en su evolución por conseguir tallos cada vez más altos. Aún no se había descubierto la estrategia de prosperar en horizontal, cosa que harían mucho más tarde las gramíneas en sus inmensas praderas. En el Devónico la tierra firme era la gran oportunidad: hacía tiempo que la capa de ozono se había formado y los rayos ultravioleta no constituían ya ninguna amenaza. La luz del sol se podía tocar ya con las manos, y las plantas fueron a por ella.
La competencia entre plantas vecinas en busca de la luz obligaba a “bombear” los nutrientes cada vez más alto, por lo que las soluciones que favorecían este bombeo o incluso el intercambio de gases en las alturas fueron las que prosperaban a través de un camino lento pero continuo. Se desarrollaron las plantas vasculares apoyadas en el perfeccionamiento de los conductos internos y de los estomas superficiales.

Evolución de las plantas, pasando desde los más ancestrales clorofitos a los traqueofitos, posteriormente a los pteridofitos, para llegar al Devónico
Ya conocimos a la riniafita Cooksonia en el Silúrico, con sus esporangios en forma de campanillas al viento, que continuó con éxito a lo largo del Devónico. Podemos considerarla casi la fundadora de la familia de los psilofitos -las plantas “φιτό”, desguarnecidas “ψιλός”-. Estas plantas, que alcanzaban una altura de 60 centímetros, carecían de raíces y casi de hojas, las cuales en todo caso eran pequeñas y parecidas a escamas, expansiones de la cutícula de sus ramillas. De todas formas tenían un pie común en forma de un tallo rastrero horizontal de donde partían vástagos ramificados, verticales y verdes.

Psilofito (a partir de BIODIDAC, términos de uso)
Otra planta de la misma familia aunque más desarrollada fue la Rhynia, de la que se disponen muchas evidencias fósiles. Presentaba unos esporangios más desarrollados y un rizoma bajo tierra que parece ser estaba asociado a algunos hongos. Esta circunstancia hace pensar que en la conquista de la tierra por las plantas tuvo mucho que ver esta relación simbiótica.

Representación de Rhynia y un corte de su tallo en donde se observan los canales vasculares internos (Wikimedia y wikimedia, GNU FDL 1.2 y Cc BY-SA 2.0 UK: England & Wales )
Los fósiles que hemos encontrado de finales del Devónico nos muestran restos de troncos y hojas, por lo que podemos imaginarnos grandes extensiones de formas arbóreas y bosques. Esta rápida aparición de variados grupos de plantas y formas de crecimiento se la conoce como la “Explosión Devónica”. Y en esta aventura tuvieron parte fundamental la utilización de nuevos polímeros como la lignina o la celulosa, que surgieron a partir de una pérdida de agua por polisacáridos y que aportaron rigidez, flexibilidad y soporte a estos organismos.
Fue una solución de ingeniería mecánica y de resistencia de materiales que resolvió un problema con la ayuda de estos polímeros, apoyados a su vez por la colaboración de la tensión superficial, que impulsaba el movimiento de líquidos por el interior de los cuerpos de las plantas venciendo a la fuerza de la gravedad.
También aparecieron otros problemas que exigían soluciones, en este caso de ingeniería bioquímica. La “irracional” e ineludible batalla por conseguir ser los primeros en llegar a los rayos de sol llevó al “gigantismo”, desde las pequeñas plantas hepáticas a los encumbrados árboles. Era evidente que la mayor altura y complexión debía ser un handicap a la hora de mantenerse erguido, por lo que poco a poco necesitaron enraizarse sólidamente en la tierra. Pero aún más crítico debió ser el que este mayor volumen corporal suponía más necesidad de nutrientes, más necesidades de agua, más necesidades de energía y más necesidades de gases atmosféricos, es decir llevaba consigo un metabolismo más ávido. En una primera instancia, el incremento en los intercambios de las moléculas necesarias para la vida y el mayor aprovechamiento de la luz solar vinieron a través de cambios evolutivos en raíces, hojas y estomas. Sin embargo, el “gigantismo” iba haciendo su labor callada hacia otra modificación menos perceptible: al crecer la masa corporal su volumen se incrementaba con el cubo del crecimiento, mientras que la sección de las redes tubulares por donde se distribuyen los nutrientes, la sección de tronco y ramas, lo hacía con el cuadrado del crecimiento. El resultado lógico debería haber sido el colapso del metabolismo por las limitaciones de abastecimiento. Pero la realidad nos demuestra lo contrario: el metabolismo no colapsó sino que se atenuó, el organismo aprendió a vivir más relajadamente, más despacio, cosa que sucederá más tarde también con los animales que agigantaron sus formas. Quizás esta circunstancia fue la que provocó en los árboles una mayor longevidad y estabilidad vital.

Imagen tomada por un microscopio electrónico de un estoma de una planta (Wikimedia, GNU FDL 1.2)
Pero con todo lo anterior no se acabaron los problemas que conllevaba la altura. Se planteaban también nuevos retos desde el área de la reproducción. Recordemos que las pequeñas briofitas vivían junto al agua, a su amparo. Y se aprovechaban de esta humedad para que los gametos generados por los tallos masculinos “nadaran” hasta encontrar un tallo femenino, allí ascender y fertilizar el correspondiente óvulo. El resultado sería un nuevo tallo con el esporangio repleto de esporas, embriones con espoleta de retardo. Cuando las esporas estaban maduras se dejaban libres y ahí donde llegaban generaban un nuevo tallo masculino o femenino que cerrarían el ciclo de reproducción. Era un poco complicado, pero eficaz. En todo este proceso el camino de los gametos masculinos era corto y la vía húmeda ayudaba. Pero al ir creciendo, esta aventura era como subir en bicicleta al más complicado puerto de montaña. Así que se optó por diferenciar los tamaños: las plantas en su fase generadora de gametos seguirían pequeñas, mientras que en la fase generadora de esporas desarrollarían altura. Pero esto era muy arriesgado en un mundo donde los animales terrestres comenzaban a campar por sus anchas, alimentándose muchas veces con los jugosos tallos verdes donde estaban las células sexuales. Así que la naturaleza seleccionó otro camino de éxito uniendo la fase sexuada y la asexuada, quedando los pequeños gametofitos incrustados en el alto esporangio: iban a llegar las plantas con semilla.
El proceso de transición fue progresivo, partiendo de unas plantas, como los musgos, en las que la fase haploide, de gametos masculinos y femeninos, era la más persistente mientras que la diploide, de embriones –esporas- la más corta en el tiempo. Se pasó después a otro tipo de plantas en las que las dos fases no eran claramente discernibles, se alternaban las generaciones de gametofitos dominantes con las de esporofitos dominantes. Le siguió una época de incertidumbre que culminó con la aparición de las plantas con semilla, en las que los gametofitos son residuales. Así en las gimnospermas, que triunfarán en el Carbonífero, los gametofitos son parte del esporofito o semilla, y en las más evolucionadas plantas con flor, las angiospermas del Cretácico, los gametofitos son unas simples células, siendo la triunfadora el esporofito que ya no es tal sino una semilla.
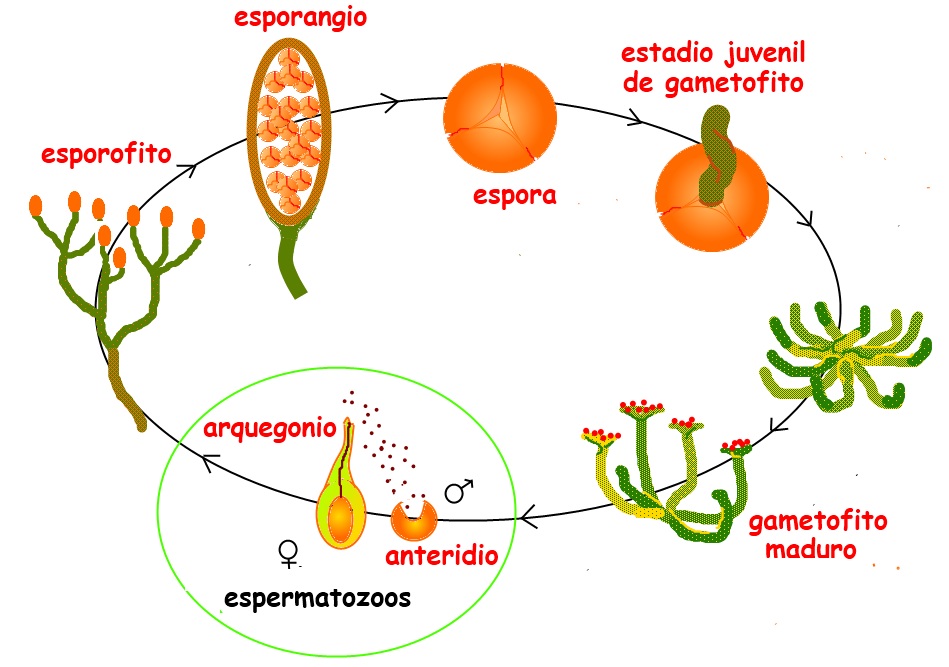
Ciclo reproductor de las Rhynias
La semilla es un embrión equivalente a una espora joven, con un tejido nutritivo asociado rodeado de una cubierta de protección. A fin de cuentas se asemeja a algo así como a un huevo animal fecundado que hubiera quedado en estado latente, a la espera de condiciones favorables para iniciar la germinación y desarrollo.
Las primeras plantas con semillas datan de hace unos 370 millones de años. El fósil más antiguo que se conoce de este grupo es Elkinsia polymorpha y consiste en unas ramitas con semillas. Otro fósil un poco posterior es Archaeosperma arnoldii, en el que se ven cuatro óvulos rodeados por unos apéndices alargados que constituyen una especie de cúpula. Estos antecesores presentaban las semillas directamente sobre las ramas, y no disponían de estructuras especializadas a su alrededor: nada que se pareciese a una flor.

A la izquierda Archaeosperma arnoldii y a la derecha Elkinsia polymorpha (Imagen: Pettitt & Beck, 1968, CNRS y Jardín Botánico de Córdoba, fair use)
Pero en realidad las plantas con semilla son patrimonio del siguiente periodo, el Carbonífero, así que vayamos a los verdaderos reyes del Devónico.
Ya hemos hablado de las riniafitas. Podemos hablar también de los licopodiofitos -que se desarrollarían en todo su esplendor durante el siguiente periodo, el Carbonífero-, de los pteridofitos ancestros de los helechos, de los que hay una gran evidencia fósil de sus troncos y hojas, o de las progimnospermas antecesoras de las modernas plantas con semilla, cuyo espécimen mejor conocido es el Archaeopteris, con 20 o 30 metros de altura, un árbol fósil con hojas de tipo helecho encontrado en todo el mundo desde el Devónico Superior al Carbonífero Inferior. No hay que confundir a este árbol con el Archaeopteryx, un animal considerado la primera ave, al cual visitaremos en la correspondiente entrada.

Representación y fósil de Archaeopteris hibernica (Imagen: Dennis C. Murphy y wikimedia, dominio público)
En un lugar próximo a Nueva York, con una historia que data de hace 380 millones de años, se encuentra el yacimiento de Gilboa. En él se han podido observar restos muy completos de un bosque que se había desarrollado en una región costera pantanosa inundada, con clima tropical, llena de grandes Eospermatopteris que recuerdan en la forma al bambú actual -aunque no está emparentado-, y con un sistema de raíces poco profundo que se extendía en todas direcciones. Ello favorecía el arraigo de otras plantas, como los Aneurophytaleans, que poblaban los suelos del bosque como los modernos helechos, formando una asociación ecobiológica con el resto de árboles, en cuyas espesas redes de raíces se apoyaban para trepar hasta el dosel del bosque de manera similar a como lo hacen algunas lianas tropicales en la actualidad.

Reconstrucción de cómo era el bosque devónico de Gilboa con los altos Eospermatopteris y las trepadoras Aneurophytaleans. En la esquina inferior izquierda asoma una Lycopsida (Ilustración: Frank Mannolini, New York State Museum, fair use)
En el mismo emplazamiento se han encontrado ejemplares arborescentes de Lycopsida, aunque son menos abundantes que las dos especies anteriormente mencionadas. Tenían unos troncos de 11 centímetros de grosor y 4 metros de altura. Estos vegetales constituyen un grupo muy antiguo de plantas vasculares sin semilla que están emparentados con los modernos licopodios. Más tarde, durante el Carbonífero, estas plantas contribuirían a la formación de abundantes masas de carbón.
Los árboles caducifolios producirían cantidades considerables de hojas que al pudrirse debían generar unos detritos muy nutritivos. El nuevo escenario provocado por este tipo de plantas tuvo que introducir muchas oportunidades en el desarrollo de la biosfera. Por un lado se incrementaban las fuentes de alimentación utilizables por los organismos heterótrofos, minúsculos artrópodos y hongos. Por otro lado aparecían otros y variados beneficios, como que la cobertura arbórea preservara de las radiaciones solares ampliándose así los ambientes húmedos, o como que el ambiente creado suponía una oportunidad para ocultarse a la vista de otros animales, ya fueran predadores o predados. Todo ello favoreció e impulsó la coevolución entre animales y plantas, entre otras la relación alimentación/transmisión de esporas/semillas. Se había abierto un inmenso nicho nuevo del que rápidamente se aprovechó la Vida. El potente océano dejó definitivamente de ser la única cuna para los organismos vivos.
Los hongos
En el yacimiento escocés de Rhynie Chert, datado en hace 410 millones de años, posee un registro fósil de hongos, en particular los más antiguos del grupo de los quitridios y de los zygomicetos -416 millones de años-. En este yacimiento se han localizado ejemplares de hongos parásitos de plantas, de forma que se pueden observar especímenes de vegetales colonizados por hifas de hongo. También muchos ejemplares presentan relaciones de saprofitismo (modalidad de nutrición a partir de los restos de otros organismos) similares a las actuales. Existen también ejemplos de hongos parásitos de otros hongos. Asimismo se ha encontrado un hongo que formaba micorrizas en las raíces aéreas y subterráneas de las plantas, como también a varios hongos zygomicetos asociados a cianobacterias en estructuras similares a líquenes. Es decir, parasitismo, saprofitismo, simbiosis de tipo liquen o microrriza… el abanico completo de la ecología fúngica actual existía ya hace más de 400 millones de años.
Vamos a dejar a los hongos en un paisaje que realmente había cambiado drásticamente a lo largo de este periodo. La tierra estaba repleta de plantas, desde las más humildes junto a los arroyos hasta las que formaban majestuosos bosques de árboles con una copa tipo palmera y hojas como helechos. Las tierras estaban asentándose gracias a sus raíces. Por debajo de estos colosos ya no sólo se oía el rumor de las aguas o el deslizarse del polvo movido por el viento, sino que de vez en cuando parece escucharse un ligero crepitar de hojas secas, un balanceo de una ramita, un tímido avanzar de un insecto, atento a conseguir comida y no ser comido. Le gusta la materia vegetal y a lo mejor confiaba en que los hongos empezaran la fermentación para aligerar su digestión. Era de esperar que los primeros hexápodos que en los mares ramoneaban algas, al salir del agua siguieran con sus hábitos alimenticios. Pero no estaban solos. Al igual que sucedía en las pantanosas costas, otros artrópodos, como pudieron ser los escorpiones marinos, les seguían la pista. El silencio tenía las horas contadas, la lucha por la vida se encargó de ello: una araña Trigonotarbida, tras una espera paciente, acababa de enrollar en sus sedas a un pequeño insecto Rhyniella. Debía alimentar a sus crías y no temía a nadie en tierra. Pero no sabía que también su época de dominio tenía los días contados…
Aunque las aventuras de los insectos es otra historia. Y la vamos a contar en la entrada siguiente, en la que también nos adentraremos en la vida de uno de sus principales predadores y actores del Devónico: los peces.

The La Biografía de la Vida 30. El devónico: desarrollo de las plantas by , unless otherwise expressly stated, is licensed under a Creative Commons Attribution-Noncommercial-No Derivative Works 2.5 Spain License.


{kind=link}
{kind=link}
{kind=link}
Escribe un comentario