
Nos habíamos quedado en la entrada anterior de esta serie en el momento en que la señal visual, que se había iniciado en una célula fotorreceptora de la retina, había conseguido llegar en forma de potencial de acción, a través de una cadena formada por dos neuronas consecutivas, una [n1] en la misma retina y la segunda [n2] en el tálamo, hasta la corteza visual primaria V1, situada en la parte trasera del lóbulo occipital del neocórtex.
Es curioso el constatar que las proyecciones de estos axones que salen del tálamo y llegan a la corteza visual V1 lo hacen físicamente de tal forma que dibujan sobre esta corteza un mapa invertido –arriba/abajo, derecha/izquierda- de la realidad exterior que impresionó la retina. No en vano el ojo, y lo que hay inmediatamente detrás, trabaja como una “cámara fotográfica”. El resultado es algo similar al homúnculo representado sobre la corteza somatosensorial -del que ya hablamos en otra entrada- o sobre la corteza motora, aunque en este caso no es espacialmente figurativo, sino de asignación espacial de propiedades de la señal. Pero sigamos adelante.
Habíamos determinado que llegaba una información muy definida de acuerdo a determinadas etiquetas: lado del escenario visto, ojo de procedencia, color e intensidad luminosa, detalle de la vía Parvo frente a generalidad de la vía Magno y todo ello de acuerdo a un mapa retiniano de puntos o pequeños círculos, que llamamos campos receptivos, manifiestamente sensibles a los contrastes luminosos más que a la intensidad puntual en sí.
Toda esta información llega a las columnas que conforman la corteza visual primaria V1, en donde se lleva a cabo un procesamiento de un nivel superior. Numerosos experimentos demuestran que el matiz de pequeñas áreas circulares de sensibilidad que comenzamos a ver en la retina se enriquece en la corteza. Todo sugiere que en el circuito visual cortical hay una organización por columnas, donde todas las neuronas de la misma columna tienden a responder uniformemente al mismo estímulo, y que el conjunto de las columnas parece también estar organizado de una forma parecida, de manera que zonas vecinas de la corteza visual tienen tendencia a responder a características similares del estímulo visual, conformándose así una especie de archipiélago neuronal de intereses funcionales. Aún más: el patrón de tal como están dispuestas físicamente en la corteza visual estas zonas coordinadas hace que el matiz particular que gestiona cada una se vaya modificando gradualmente si nos movemos de una zona a la vecina. De forma que, al final, la corteza visual presenta un patrón semejante a una conjunción de columnas dispuestas más o menos de forma circular como las hojas de una margarita, en la que se van repartiendo por sectores grupos de neuronas que responden al mismo estímulo, respuesta que va variando suavemente a medida que vamos dando la vuelta alrededor del conjunto. Y lo más curioso es que los estímulos de los que estamos hablando no reproducen como podríamos imaginar aquellos contrastes puntuales que vienen de la retina, sino que reproducen estímulos lineales, cada uno con sus preferencias de orientación –se entiende ahora la disposición circular, que permite jugar por sectores especializados con los 360º- y de dirección y sentido de movimiento de la “barra” estímulo –dentro de cada sector especializado en una orientación hay neuronas que prefieren procesar el movimiento del estímulo en un sentido y vecinas de ellas que prefieren el sentido contrario-.

Imágenes reales de la distribución neuronal en la corteza visual V1 por preferencias en la respuesta a la orientación. Arriba, resultado de un par de experiencias en las que las partes más oscuras corresponden a las neuronas que se activan ante un estímulo “barra” con la orientación indicada. Abajo composición cromática obtenida a partir de las experiencias anteriores. Cada color representa una orientación según el código inclinación/color situado en el centro de la figura. En la ampliación de la derecha las flechas apuntan al centro de lo que hemos visualizado en el texto como una “margarita”. En una la variación en la percepción de la orientación sigue un giro en un sentido, mientras que en la otra lo hace en sentido contrario (Imagen, fair use)
Esta información más rica que se está procesando en la corteza visual primaria V1 va a seguir su camino hacia circuitos más complejos y más profundos en el corazón de las funciones cerebrales. Efectivamente, a través del tejido cortical la corteza primaria V1 se proyecta a las cortezas visuales de asociación vecinas, que podemos imaginar como dispuestas prácticamente en anillos concéntricos alrededor de V1. Se están realizando estudios intentando definir exactamente cuántas de estas áreas hay, pero aún no están las cosas claras, aunque sin lugar a dudas encontramos a medida que nos movemos por ellas una representación cada vez más integrada acerca de particulares aspectos del mundo visual. Cada una de estas nuevas cortezas visuales parece que están encargadas de un aspecto concreto de la percepción visual: las formas y figuras, tal como se mueven o varían, el color, la velocidad, la dirección del movimiento… aspectos que tenemos marcados de forma gráfica en la última imagen de esta entrada.

Distribución jerárquica de las cortezas visuales. V1 es la corteza que primero recibe la señales “brutas” que provienen de la retina (Imagen de la red, fair use)
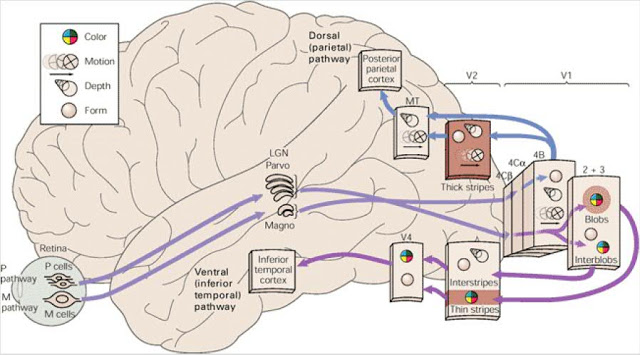
El primer paso tras el amasado en “barras” se realiza en la corteza V1, y la información así preparada pasa a la corteza V2, la cual está organizada por áreas especializadas, cada una de ellas en una característica concreta del campo visual, como lo son el color, la forma, el movimiento, la orientación… Es decir, se pasa del manejo integrado y unitario de los diversos aspectos que incorpora la información del campo visual -lugar, lateralidad, contraste lumínico…- que se realizaba en las “columnas” de la corteza V1, a otro manejo en el que se disgregan e integran los datos anteriores de otra manera, según matices, para convertirlos en cualidades específicas de la visión que se gestionan como realidades independientes. A partir de este mundo tan variado de aspectos parciales de la percepción de la visión, parecen conformarse dos grandes vías a medida que se van integrando procesos en las cortezas asociativas. La primera, que se dirige hacia el lóbulo parietal, se conoce como la vía del “cómo”, y la segunda, que se dirige al lóbulo temporal, conocida como la vía del “qué”. El “cómo” responde a dónde se encuentran los objetos y cuáles son las características de sus movimientos. Originada en la corteza visual V5, hace un mapa del mundo exterior mediante el que puede realizar un análisis de los patrones de movimiento vistos: en qué posición está el coche, a que velocidad se acerca, el posible punto de colisión conmigo…. El “qué”, originado en la corteza especializada V4, se refiere a ¿qué es lo que estoy viendo? Tamaño, color y forma. A la identidad de una fruta, su color es amarillo, tiene una textura lisa y brillante, es alargado y no muy grande, se trata de un plátano… quizás conceptos relacionados casi con la semántica -función cerebral que parece se procesa también en el lóbulo temporal-. En los humanos esta vía nos ayuda también a reconocer las caras y la identidad de las personas a las que estamos viendo. Podemos apreciar mejor lo dicho acerca de los caminos de ambas vías -procesos especializados- en la imagen siguiente.

Esquema general de las vías de la percepción de la vista, desde la retina hasta las cortezas más profundas, con indicación de la característica visual que se gestiona en cada zona. Se han representado las dos vías parvo y magno equivalentes a las del “qué” y el “cómo”. En las cortezas V1 y V2 se ha dado el detalle gestionado en sus capas neuronales. Los “blobs” son lo que hemos figurado en el texto como “margaritas” (Imagen de la red, fair use)
Como conclusión general podríamos decir que las áreas de la corteza visual gestionan un aspecto específico de la imagen captada en la retina, contribuyendo de forma directa, aunque incompleta, al hecho último de la percepción visual. Es evidente que entre todos estos módulos debe haber una red potente de conexiones que permita integrar los distintos aspectos gestionados, que no sólo se llevan a cabo en lugares físicos distintos, sino que también a lo largo de periodos temporales distintos. Por ejemplo, percibimos el color 40 mseg antes que la forma, y ésta 80 mseg antes de que percibamos el movimiento. Y parece que el cerebro lo hace en base a datos muy elementales, como pueden ser las barras luminosas con una determinada orientación, que luego integra sin que sepamos muy bien como lo hace. Incluso este software integrador puede ser distinto en cada una de las cortezas visuales. Aunque sí estamos seguros de que en ello no sólo interviene la máquina que hemos heredado durante nuestro desarrollo ontológico, producto de nuestra carga genética, sino también que su respuesta está también muy influida por la experiencia real, por factores epigenéticos. Es significativo que si privamos de la visión a algunos mamíferos -y nosotros lo somos- durante las primeras semanas de vida, este animal adquirirá con seguridad algún tipo de ceguera funcional. La verdad es que desconocemos cuál pueda ser el módulo integrador unitario del cerebro dedicado a la vista, si es que existe uno solo.
Las opiniones[1] del prestigioso neurobiólogo turco Semir Zeki ayudan a dar un poco de luz a este nudo gordiano. Él fue el que descubrió la funcionalidad de algunas de las cortezas visuales secundarias como la V4. De entrada, el cerebro, aunque es un órgano muy potente, es también limitado. Y por tanto deberá aliviar su capacidad de memoria y computación trabajando selectivamente en base a ciertas características constantes y generales que existen en el mundo externo. Y esto quizás lo hace en base a su capacidad de generar abstracciones -seleccionar de lo general lo más simple, propiedades y relaciones que se repiten-, para luego, basado en estas abstracciones sencillas y universales, inducir lo que Zeki llama ideales, que no dejan de ser más que esquemas generales de gestión de información, como pueden ser “esto es un perro” o “esto es bello“. En sus propias palabras: “Yo defino el ideal… de un objeto como el registro que el cerebro hace de todos los objetos que ha visualizado [color, forma, tamaño, temporalidad...], siendo éste dependiente de una multiplicidad de percepciones concretas, y a partir del cual el cerebro es capaz de sintetizar un ideal”. Y sigue: “…la abstracción y el idealismo implican -mediante un proceso neural que nos es desconocido- la selección de rasgos o características (aquellas más importantes para el sujeto), el rechazo de otros y la síntesis de los que son seleccionados en una nueva entidad ligada a las experiencias individuales al tiempo que indiferente a cualquier experiencia concreta“. El cerebro llegaría a producirnos la percepción visual -y podemos generalizar a cualquier percepción- en base a encontrar en lo que ve las abstracciones e ideales implícitos -cosa que se haría en las diversas cortezas V-, para luego, en proceso asociativo, compararlas con sus patrones de abstracciones e ideales internos, conseguidos a través de la experiencia, y así determinar lo que debe proponernos como mejor opción de percepción de lo que vemos.
Con eso damos por terminado nuestro propósito, que no era otro que describir cuales eran los caminos de entrada de la información visual exterior y seguirlos hasta las zonas de procesamiento complejo cerebral. Hemos visto cómo la información que al final nos produce la percepción de “ver”, entra a través de una cadena de potenciales de acción todos muy semejantes y, lo que es más importante, a través de dos vías paralelas, cada una de ellas conductora de matices particulares y esenciales, que desembocan también en zonas corticales, en consonancia, muy específicas. Son los grupos de neuronas que las forman, trabajando en conjunto, los que hacen posible los variados y ricos matices que creemos apreciar en nuestras percepciones visuales. Es, pues, el cerebro el que inventa lo que creemos y sentimos ver. ¿Homotético de lo que hay afuera? ¡Quién lo sabe! Realmente debe representar a lo que hay fuera, pero ¿qué es lo que hay fuera sino unas interacciones de los campos electromagnéticos que al excitarse transmutan su energía en lo que creemos partículas, que llamamos fotones? Ya en el siglo XVIII el filósofo napolitano Giambattista Vico escribía: “Si los sentidos son capacidades activas, de ahí se deduce que nosotros creamos los colores al ver, los gustos al gustar y los tonos al oír, así como el frío y el calor al tocar”.
En la siguiente entrada nos adentraremos en los caminos neuronales sensitivos de otro viejo sentido: el oído.
- Texto aquí [↩]

The Los sistemas receptores 11: La visión III. Procesos superiores. by , unless otherwise expressly stated, is licensed under a Creative Commons Attribution-Noncommercial-No Derivative Works 2.5 Spain License.


{kind=link}
Escribe un comentario