
Atención: NOTA IMPORTANTE
Tengo que advertir de que en esta entrada y en alguna de las que la seguirán voy a obviar hasta cierto punto el criterio de organizar el devenir de los sucesos de forma cronológica, siguiendo el paso del discurrir geológico. Soy consciente de que el cambio puede llevar quizás a confusión al lector asiduo de la serie. La explicación es sencilla: aprovechando el impulso que nos da el esplendor de las radiaciones del Cámbrico creo que viene como anillo al dedo abrir en este momento un serie de paréntesis en los que se vaya analizando paso a paso, independientemente de su duración temporal, cada uno de los diversos caminos que siguió la biota en pos de la rica complejidad de formas que surgieron en el entorno de este periodo. Es obligado poner el foco sobre diferentes sucesos evolutivos de gran trascendencia de forma que podamos entenderlos y ver, por ejemplo, cómo pudieron aparecer la simetría bilateral, la notocorda, el aparato locomotor o el ojo, entre otros.
La claridad en el desarrollo argumental de cada uno de estos importantísimos aspectos de la evolución de la Vida exige a su vez una vida cronológica propia para cada uno de ellos. Así que nos moveremos desde el Precámbrico y a lo largo del Fanerozoico siguiendo el desarrollo evolutivo de cada uno de ellos, por lo que la sucesión de periodos se irá repitiendo al compás de cada nuevo monográfico. Intentaré hacer un breve recuerdo de esta advertencia al principio de cada una de las entradas afectadas.
En la entrada anterior de esta serie sobre la Biografía de la Vida comentamos la expansión acelerada de nuevas formas de seres vivos en lo que se conoce como la explosión del Cámbrico. En aquellos momentos, hace unos 540 millones de años, se iniciaron los principales filos de animales que hoy podemos observar: artrópodos, braquiópodos, moluscos, anélidos o vertebrados. La evidencia fósil durante el Cámbrico es clara para todos ellos.
Continuamos hoy hablando de cómo pudieron iniciar la andadura cada uno de ellos, desde el Cámbrico hasta periodos posteriores. Y como elemento que ayude a seguir la saga, repetiremos aquí el árbol con el que cerrábamos la entrada anterior.

Caminos evolutivos de los especímenes de la fauna del Cámbrico. La imagen del ancestro común corresponde a una Planaria actual (a partir de imágenes de wikimedia)
Un preludio dedicado al gusano platelminto
En el esquema de la figura anterior vemos una raíz única, personalizada en un gusano platelminto. Por su posición de posible padre común de los principales filos de los metazoos actuales, bien se merece que le dediquemos una breve genealogía.
El biólogo Robert D. Barnes postulaba tres posibles vías por las que se llegó al primer organismo del Reino Animalia. Una de ellas parte de un animal ciliado, como el paramecio, que duplicó multitud de núcleos en su citotoplasma y que con posterioridad los aisló mediante nuevas membranas. El resultado sería un organismo pluricelular. Dado que muchos de los ciliados actuales presentan simetría bilateral, bien pudo ser que el ancestral ciliado que pasó a multicelular adoptara esta simetría, transformándose en un platelminto (teoría sincitial).
La segunda teoría se apoya en los flagelados unicelulares. En una entrada posterior nos encontraremos con el caso del alga Volvox, y veremos cómo estos flagelados se agrupan formando unas esferas huecas. Con el tiempo estas esferas pudieron sufrir una invaginación conformando una línea de organismo de tipo medusa, para después rellenar su cavidad interna con nuevas células de sus paredes, transformándose a la larga en un platelminto (teoría colonial).
Por fin, una tercera hipótesis es más generalista, ya que postula un paso evolutivo no especificado que transformó a algún ser unicelular en un platelminto.
Lo anterior nos lleva a lo que ya hablábamos en la entrada anterior al analizar el árbol filogenético del Proterozoico, en donde comentábamos cómo las plánulas y los acelomorfos podían estar en la base de los animales bilaterales, y en consecuencia en la base de los platelmintos, los primeros de ellos.

Evolución de los metazoos primitivos hacia el gusano platelminto, según las teorías de Robert D. Barnes
Sigamos ahora con el hilo principal atendiendo al filo de los artrópodos.
El camino de los artrópodos
Más allá de 536 millones de años atrás, un cuerpo cilíndrico con unos apéndices que se pueden clasificar como patas buscaba su vida protegido entre las colonias de esponjas. Unas patitas que básicamente eran una funda de músculos llenas de un líquido, que podríamos asemejar funcionalmente a la sangre, y que servían para moverse: los lobopodos. El primer fósil lobopodo se llama Aysheaia pedunculata, y tenía espinas en sus patas que le servían para recolectar comida de los tapices bacterianos. A menudo disponían de numerosas placas blindadas que cubrían todo el cuerpo y la cabeza. Pero como eso les dificultaba la defensa, desarrollaron espinas. No se parecía a un artrópodo, pero iba camino de ello.

Reconstrucción de Aysheaia pedunculata (Wikimedia, GFDL 1.2)
De los Lobopodia pasamos a Diania cactiformis. Se trata del primer ejemplar con patas articuladas típicas de los artrópodos. Es el primer Lobopodia con un exoesqueleto simple. La criatura que vivió hace unos 520 millones de años medía 6 centímetros de largo. El fósil no muestra signos de la existencia de ojos o mandíbulas, pero tiene 10 pares de patas articuladas. Se cree que estas patas tenían una superficie dura no muy diferente de las que hoy recubren los miembros de crustáceos o insectos. Se alimentaba del plancton y del detritus orgánico del fondo marino, y la armadura de sus patas probablemente se desarrolló como protección frente a los primeros depredadores.

Recreación de Diania cactiformis (Wikimedia, CC BY-SA 3.0)
Y del cactiformis al trilobites y la familia artrópoda. Evidentemente el tránsito se hizo a través de varios escalones, pasando por la exoesqueletización del cuerpo y la aparición de los ojos. Del trilobites se hablará un poco más en la descripción del Ordovícico.

Fósil del trilobites Phacops rana (Wikimedia, CC BY-SA 3.0)
Hace unos 515 millones de años ya se observan en los artrópodos unas morfologías semejantes a la de una gamba de hoy en día, es decir, bivalva en la parte dorsal y cuerpos segmentados acabados en un par de aletas. Algunos de sus fósiles nos han dado una gran información sobre la reproducción de estos animales. Se trataría de Kunmingella douvillei uno de cuyos especímenes, que procede del yacimiento chino de Maotianshan Shale, lleva en su interior huevos aún no fecundados. Un poco posterior es Waptia fieldensis especie de la que cinco de sus fósiles encontrados[1] en Burges Shale, datados en 508 millones de años, portaban, éstos sí, huevos con embriones protegidos bajo su bivalva, lo que indica que esta estructura facilitaba su transporte, permitiendo el cuidado de las crías. Seguramente son las madres más antiguas de las que tenemos conocimiento.
Emparentado con los artrópodos vivió el extinguido Anomalocaris, la “gamba anómala”, el rey de la depredación del momento, hace de 525 a 510 millones de años. Siempre se ha creído que su visión debía aterrorizar. Podía medir hasta un metro de longitud y se movía con gran agilidad gracias a los numerosos lóbulos laterales y a los tres que conformaban su cola. El cuerpo articulado poseía un exoesqueleto orgánico. Aunque realmente lo que debía impresionar era la visión de su cabeza, con dos grandes apéndices llenos de espinas con los que atrapaba a sus presas y se las llevaba a la boca donde los trituraba, ya que en ella disponía de un montón de láminas aserradas.
Los últimos estudios de sus fósiles realizados en 2009 por el paleontólogo del Museo de la Naturaleza y la Ciencia de Denver, James W. Hagadorn, parecen dar más luz a sus hábitos alimentarios, ya que determinan que la disposición de su boca no permitiría ejercer una fuerza suficiente como para romper caparazones duros como el de los trilobites, y que quizás tuvieran una forma de alimentarse más generalista con un amplio abanico de hábitos alimentarios, desde depredador acechando a la emboscada hasta realizar una criba de sedimentos en busca de carroña o diminutos restos orgánicos.
El Anomalocaris lo tendría todo controlado mediante un par de ojos compuestos, tipo yunque, que le daban una visión clara de la que se aprovechaba para cazar con precisión en las bien iluminadas aguas de los mares cámbricos. Para procesar esa información tenía que haber desarrollado ya un cerebro en consonancia. Esta hipótesis no resulta sorprendente, puesto que hay pruebas moleculares que indican que las estructuras clave del cerebro humano datan de hace más de 600 millones de años.
No obstante, no todas las especies de este artrópodo eran tan terroríficas. Recientemente (2014) se ha comunicado el descubrimiento[2] en un yacimiento de Groenlandia de un “especial” anomalocárido de hace 520 millones de años, Tamisiocaris borealis. Sus apéndices frontales habían desarrollado la forma de una red, con la que efectuaba algo así como una pesca de arrastre, atrapando por filtración plancton de hasta medio milímetro de tamaño. Se cree que esta especie adoptó esta solución para su alimentación en respuesta a la competencia que suponía el entorno de gran presión depredatoria del Cámbrico, lo que les obligó a cambiar de nicho ecológico.

Imagen artística de Anomalocaris saron depredando trilobites (Wikimedia, GFDL 1.2)
Habrá que esperar hasta la frontera del Silúrico con el Devónico, unos cien millones de años después (416 mm aa), para que los artrópodos dejaran el agua y conquistaran la tierra, abriendo el nuevo mundo de los insectos, arañas, escorpiones y miriápodos.
La siguiente figura indica la correlación filogenética entre los diversos subfilos de los artrópodos, en donde se muestran los momentos de las divergencias entre ellos. Está basada en estudios[3] realizados en 2009 por el biólogo Davide Pisani sobre las secuencias de genes nucleares y mitocondriales de estos animales. Los resultados son discrepantes en cuanto al punto de partida temporal de este filo con los que dimos al hablar del Proterozoico, cuando presentábamos la cronología filogenética basada en la información del libro “The ancestor’s tale” (2004) de Richard Dawkins, en donde el punto de divergencia de los artrópodos se situaba unos cien millones de años más tarde. El esquema de Pisani amplía la información, pudiéndose observar el parentesco tan próximo entre crustáceos e insectos, que quedan enlazados por un hexápodo ancestral que apareció por evolución de un crustáceo en un ambiente de agua dulce cuando aún no se había conquistado el medio terrestre.

Aparición y diversificación de los artrópodos (de acuerdo a los estudios de Davide Pisani, 2009)
El camino de los braquiópodos
Por la misma época en que los artrópodos comenzaban su carrera de éxito, hace unos 555 millones de años habitaba el fondo marino un extraño animal ovalado, bilateral, que al igual que una babosa se encogía y alargaba para moverse en una dirección: el Kimberella. Sus restos fósiles se han encontrado en el yacimiento australiano de Ediacara y con posterioridad en la región rusa del Mar Blanco. Estaba recubierto por un tegumento no mineral, firme pero flexible, que no le impedía el movimiento, formado por una especie de pequeñas escamas que se hacían más duras a medida que el animal crecía, y le rodeaba una especie de volante que haría las funciones de agallas rudimentarias.

Representación artística de Kimberella (Wikimedia, CC BY-SA 3.0)
Más tarde, hace unos 525 millones de años, el Kimberella convivía en el mar con otros seres, quizás uno de ellos el Halkieria, un animal con cuerpo plano y simetría bilateral, con un “vientre” liso y pegajoso como el de una babosa. Se movía por el fondo marino mediante este vientre mucilaginoso. Su parte dorsal estaba protegida por escamas huecas ancladas a la espalda mediante un tallo, formando una especie de armadura. Y lo más curioso es que sobre la espalda, en sus dos extremos, acarreaba dos conchas parecidas a valvas, la posterior muy semejante a la de los braquiópodos del Cámbrico. Una posible explicación de por qué desarrolló esta extraña morfología es que le era sumamente útil cuando se veía atacado: se plegaba por la mitad, más o menos protegido dentro de las dos placas. Como la predación en su época era la norma, quizás la evolución le llevó a adoptar definitivamente esta forma, doblada, anclándose en un lugar. Se trata de la Halkieria evangelista.

Fósil de Halkieria evangelista en el que se aprecian claramente las dos valvas que protegían ambos extremos de su cuerpo (Wikimedia, GFDL 1.2)
Después de contemplar a Halkieria es casi inevitable el sospechar el siguiente paso. ¿No parece creíble que la cadena evolutiva “gusano Platelminto-Kimberella-Halkieria” nos deba llevar hasta los Brachiopoda? A la vista de la imagen siguiente no es difícil imaginar a un braquiópodo como un Halkieria doblado en posición defensiva.

Fósil del braquiópodo Leptaena acuticuspidata (Consejería Educación Junta de Andalucía, fair use)
Los braquiópodos se diferencian de los moluscos bivalvos en que sus dos conchas presentan una disposición de superior/inferior, unidas en la región posterior, mientras que en los moluscos son bilaterales (derecha/izquierda).
El camino de los moluscos
Ya hemos hablado del Kimberella como posible ancestro de los braquiópodos. Pero también lo fue de los moluscos, según los estudios genético-moleculares. Junto a sus fósiles se encuentran marcas muy similares a las que hacen los moluscos cuando raspan con sus rádulas, su característico aparato alimentador. Aunque en algún caso, como el de los especímenes encontrados en los yacimientos rusos del Mar Blanco, ninguno presentaba signos de poseer rádula.
También la mencionada Halkieria, ornamentada con una valva en cabeza y otra en cola, está en el camino de los moluscos a través de Viwaxia. Ambos tienen puntos en común, una base blanda tipo babosa mediante la que se desplaza, y escamas huecas con una misma estructura microscópica y ordenadas también en tres zonas. Pero Viwaxia había dado un paso más. Es el animal más antiguo que conocemos con rádula.

Representación artística del Viwaxia (Wikimedia, GNU FDL 1.2) y rádula de un molusco actual (Wikimedia, GFDL 1.2)
Más allá de estos animales se encuentra lo que pudiera ser el primer molusco, el aplacóforo. Literalmente significa “que no lleva concha”. Tiene apariencia de gusano recubierto de espinas calcáreas. Se supone que por fusión de estas espinas se llegó a la concha de los verdaderos moluscos. Bien pudiera ser que las escamas de Viwaxia pudieran sufrir un proceso semejante.

El aplacóforo Epimenia verrucosa (Wikimedia, GFDL 1.2)
El Nectocaris pteryx, un animal fósil de hace 500 millones de años, era un antepasado carnívoro de los calamares y por tanto el antecesor más antiguo que se conoce de los moluscos cefalópodos. Un reciente estudio revela que Nectocaris era similar a los miembros del conocido grupo moderno de los cefalópodos -que incluye pulpos, calamares, sepias y nautilus, así como los ya desaparecidos ammonites y belemnites perdidos en pasadas extinciones masivas-.

Fósil (Wikimedia, dominio público) y representación artística de Nectocaris pterix (Wikimedia, GFDL 1.2)
Los especímenes fósiles, de entre dos y cinco centímetros de largos, muestran que Nectocaris tenía forma de cometa y se aplanaba desde la cabeza a la cola. Tenía un par de ojos pedunculados y un par de tentáculos largos para atrapar las presas que consumía. Se cree que esta criatura utilizaba sus aletas laterales para nadar y, como los modernos cefalópodos, también usaba una tobera en forma de embudo para acelerarse mediante propulsión a chorro.
El camino de los anélidos
Aquí tenemos que seguir acudiendo a Viwaxia. Los poliquetos actuales son gusanos anélidos con una apariencia peluda o espinosa. Los pelos se llaman quetas y están mineralizados: la microestructura de estas quetas es idéntica a la de las escamas de Viwaxia. Y todavía hay más conexiones: Canadian, un gusano anélido de los poliquetos, cuenta con una concha quitinosa en uno de sus extremos. Sorprendentemente, un espécimen de Viwaxia presenta también en un extremo una especie de concha que le emparenta con su antecesor Halkieria. Y no sólo la relación se da en la morfología de las conchas de poliquetos con las de Viwaxia y Halkieria, sino que los análisis moleculares así parecen avalarlo.

El poliqueto actual Nereis succinea (Wikimedia, CC BY-SA 3.0)
El camino de los vertebrados
La historia de los vertebrados empieza con el invento de los cordados durante el Cámbrico. La corda, estructura que les da nombre, es una varilla que se extiende a lo largo del cuerpo y que tiene una rigidez suficiente como para proporcionarles un sostén. Se sitúa en posición dorsal con respecto al tubo digestivo.
Un grupo de investigadores del instituto médico norteamericano Howard Hughes afirma en un reciente estudio (2014)[4] que la notocorda evolutivamente procedería de estructuras musculares que habrían desarrollado animales aguas arriba del árbol filogenético cordado. Posteriormente este “fleje” muscular denominado axocorda desarrollaría un bastón central cartilaginoso, elemento que daría mayor eficiencia a la natación (tal como se observa en el moderno anfioxo).
Según Richard Dawkins, éste fue el árbol filogenético familiar.

Árbol filogenético según Richard Dawkins
Comienza la saga con unos fósiles encontrados en el yacimiento de Maotianshan Shales, en China, datado en el bajo Cámbrico de hace 525 millones de años. En él se han encontrado, entre otros, restos de animales que parecían iniciar la línea de los peces: el Haikouella, un “gusano” cartilaginoso que poseía ojos, intermedio entre los invertebrados. Se trata aún de un primitivo cordado.
El Haikouella tenía notocorda, hendiduras branquiales por las que absorbía agua y cribaba los alimentos y arcos branquiales, que son pequeñas barras cartilaginosas entre las hendiduras branquiales. Son particularmente importantes estas características ya que representan la primera evidencia de un armazón, aunque cartilaginoso, que sustentaba longitudinalmente al cuerpo y que con el tiempo derivaría hasta la columna vertebral de los vertebrados. Evidencia una estructura de hendiduras y soportes gelatinosos cuyo diseño es semejante al que presentan los peces y al que tienen los embriones de los tetrápodos durante su desarrollo inicial. Con posterioridad veremos que estas estructuras son las que conformaron los huesos, nervios y músculos de la cara y cuello de los mamíferos. Sin duda el Haikouella es el ancestro más remoto con un plan corporal precursor, y además ya similar al nuestro.
También en los yacimientos chinos de Maotianshan Shales se ha encontrado el Haikouitchtys (el mismo nombre le delata, “pez de Haikou”). Parecido a Haikouella aunque ya había iniciado el camino de la especialización encontrándose clasificado en el subfilo craniata, los craneados. Contaba por tanto con una estructura craneal, quizás el más antiguo cráneo descubierto hoy en día.
Una noticia aparecida en junio de 2014[5] nos permite expandir el campo geográfico de este tipo de animales antecesores de los vertebrados. Debían ser muy ubicuos en los mares del Cámbrico. En el otro extremo del mundo, en el yacimiento de Burgess Shale de Canadá, se encontró lo que bien pudiera ser un nuevo “nieto” del cordado Haikouella. Se trata de fósiles de Metaspriggina, datados en hace 505 millones de años, del que se ha comprobado la proximidad filogenética con Haikouitchtys. Se ha podido determinar su especial disposición de los arcos braquiales, dobles, que permite hacer un seguimiento en el tiempo de la evolución de las mandíbulas. En una entrada posterior, la número 29, analizaremos la relación entre ambas estructuras corporales.
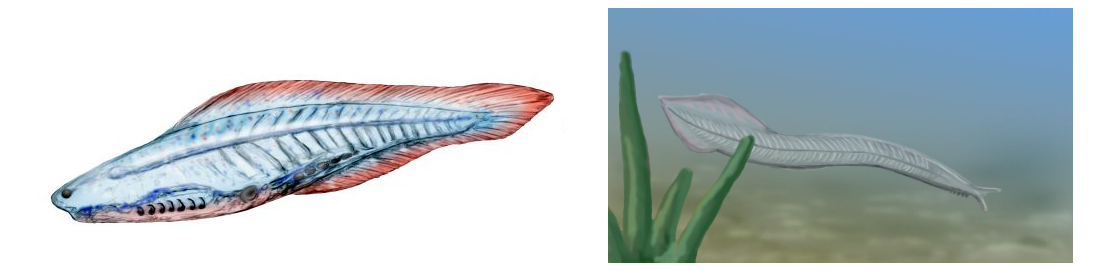
Representación artística de Haikouitchtys, izquierda, (Wikimedia, CC BY-SA 3.0) y de Pikaia gracilens, derecha, (Wikimedia, GFDL 1.2)
También del yacimiento de Burgess Shale es el fósil de Pikaia, datado también en hace unos 505 millones de años. Inicialmente se le consideró como uno de los ancestros más antiguos de los vertebrados. Esta afirmación presenta muchas dudas para los círculos científicos y parece que filogenéticamente se debe encontrar más próximo a la línea de los cefalocordados (ver figura anterior). Disponía de un rudimentario cordón “columna vertebral”, formado por cartílago junto a su médula espinal, que no habían desarrollado un engrosamiento de la corda en el cráneo a manera de encéfalo. De ahí el nombre del subfilo. En promedio medía 5 centímetros de longitud y debía nadar con facilidad sobre el fondo marino mediante movimientos ondulatorios, usando su cuerpo y una prolongación de su cola como timón.
Siempre se ha dudado sobre cuál pudo ser el siguiente paso hacia los modernos vertebrados ¿el rudimentario esqueleto óseo ancestral provendría de una variación de las estructuras dentarias o bien de protecciones externas resultantes de un endurecimiento de la piel?
La primera vía la representa el Conodonto, un cordado parecido a una anguila. No tenía mandíbulas y se piensa que se alimentaba a través de una boca succionadora muy parecida a la de las actuales lampreas. En esta boca se encontraban unas estructuras óseas semejantes a dientes. Una especie de lengua los sacaba e introducía de la boca, movimiento mediante el que posiblemente raspaba y cribaba las rocas cubiertas de algas. Incluso les servirían para atrapar a otro tipo de presas, por lo que con el tiempo estos animales se convirtieron en feroces depredadores. Estos dientes eran de fosfato de calcio y es lo que permite pensar que pudieran ser los más primitivos indicios del hueso. Aparecen hace 500 millones de años y se siguen encontrando hasta el Triásico, hace unos 200 millones de años

Representación del Conodonto y sus placas (agnatosactuales.wikispaces.com, CC BY-SA 3.0)
La segunda vía de radiación de los vertebrados es la representada por los animales con coraza protectora externa y aparece con los Ostracodermos. Ellos fueron los primeros peces y aún no habían desarrollado las mandíbulas. Sus fósiles están datados en hace 475 millones de años durante el Ordovícico. En la entrada correspondiente hablaremos más de ellos al continuar la saga de los vertebrados.
Una vez presentados los actores nos seguimos preguntando ¿La línea filogenética de los vertebrados se dispone a través de la serie Pikaia/Conodonto/Ostracodermo o bien tras Pikaia se abren en paralelo por un lado la línea de Conodonto y por otro la de Ostracodermo/resto de vertebrados?
Las últimas investigaciones parecen demostrar que el último planteamiento[6] es el correcto, de forma que en la historia evolutiva se habría dado por duplicado el episodio de generación de estructuras óseas, una en los conocodontos y otra en los primeros peces acorazados que es la que habría llegado hasta nuestros días.
Adquiriendo habilidades
Este esplendor de vida del Cámbrico se materializó en cincuenta nuevos grupos animales que en apariencia no tenían antecesores conocidos. La mayoría, con simetría bilateral que continúa hasta hoy en día, demostrando que es una habilidad ganadora.
Partíamos de una fauna del Precámbrico tardío que no había desaparecido completamente como lo había hecho la ediacara, ya que su acervo genético les dio alguna ventaja que les favoreció, seres microscópicos que habrían dado el salto ayudados por nuevas habilidades que se habían conseguido, ya desde el Precámbrico, a través de pequeños pasos. Está claro que con cada cambio se abría para el organismo que lo había desarrollado un horizonte de nuevas posibilidades, llevándolos a formas distintas y más complejas, que no necesariamente más perfectas. Simplemente una alternativa más que podía sobrevivir… o no.
Ya se ha comentado que uno de los detonantes que favorecieron la rápida diversificación bien pudo ser el cambio de estrategia a la hora de buscar alimento, actividad que se independizó del azar (comer cuando la comida se cruzaba en tu camino), para ir activamente en su busca en vez de permanecer a la espera, lo que pudo estimular una carrera armamentística entre predadores y predados. En esta carrera las nuevas habilidades eran permitidas por la selección natural. Un elemental sentido de la visión suponía una ventaja a la hora de buscar comida o cobijo, lo que diferenciaba positivamente a quien lo tuviera del vecino que no lo hubiera desarrollado. Una cuerda cartilaginosa permitía el soporte del cuerpo y reforzaba la capacidad de movimiento. Y más útiles complejidades: ¿Por qué la simetría bilateral? ¿Cómo aparece el cráneo? ¿Y el éxito de los ojos? ¿El porqué de las mandíbulas? ¿Cómo se desarrollaron las “patas”? ¿Por qué llegó la especializada segmentación corporal? ¿Cómo apareció el sistema nervioso que controlaba toda esta complejidad?
Las siguientes entradas serán como un escaparate en donde analizaremos cómo fueron los detalles que contestan a estas preguntas. La clave la tiene la mutación en el genoma particular de cada animal, cualquiera que fuese la vía que se siga, y la posterior selección natural. Pero habrá que esperar un poco, ya que antes nos toca hablar de esta última, la selección natural, cosa que haremos en la siguiente entrada.
- La reseña sobre este hallazgo la podéis encontrar en este enlace a Courrente Biology, de enero de 2016. [↩]
- La información de más detalle la encontráis en este enlace. [↩]
- Los podéis encontrar en este enlace. [↩]
- Que podéis encontrar en este enlace. [↩]
- Aquí podéis encontrar la referencia mencionada publicada en Nature en junio 2014. [↩]
- La publicación que habla de esta circunstancia la encontraréis en este enlace. [↩]

The La Biografía de la Vida 19. Los filos del Cámbrico by , unless otherwise expressly stated, is licensed under a Creative Commons Attribution-Noncommercial-No Derivative Works 2.5 Spain License.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{ 4 } Comentarios
Maravilloso y todo de algo parecido a un platelminto. No sera por que haya poca informacion, pero a mi me gustaria ver algo mas de comentarios del tipo evolucion, “resulta que a los alienigenas les encanta abducir celulas individuales pero les molesta sobre manera las que estan agrupadas en organismos aceloides, esa asquerosa baba no se puede saborear bien, por lo que adoptar esta configuracion era una ventaja para la supervivencia”, “como todo el mundo sabe los hombres lobo acuaticos solo temen a dos cosas, a la calvicie y a los ojos saltones, el Anomalocaris se aprovechaba de esto” o algo que sea verdad. Claro que no se que conocimientos de esos aspectos tenemos, ahi sutilezas que se nos escapan en especies vivas, no tengo ni idea de si eso se podra hacer en registros fosiles. Claro que ahora que me fijo creo que eso aparecera en siguientes capitulos y simplemente soy un ansioso. De todas formas aprovechar para dar felicitaciones por la serie como siempre, que ultimamente no lo hacia.
Hola Sergio B. Yo creo que esta serie que surgió con un deseo de compartir el resultado de mis lecturas con gente que tuviera parecidas inquietudes y curiosidades, no tiene más interés que el que entre todos me ayudéis, o nos ayudemos, a incrementar el conocimiento sobre este tema tan interesante. Esto más que un mundo es un multiverso. Y cada día que leo más cosas me va entrando como un canguelo por la cara que hay que tener para exponer en público la evolución de la Vida, cuando hay ya tanto escrito y tan profundo. Pero bueno, a divertirse y a compartir comentarios con amigos de la red.
Personalmente creo que la serie da una buena visión generalista de lo que pudo pasar aunque se queden en el tintero muchísimas cosas. De la mayoría de ellas no soy ni consciente y cuando me voy asomando poco a poco a algún detalle me va entrando el vértigo. Así que me alegra el ver que hay gente como tú que espera encontrar “proteína de la buena” en mis escritos. Poco a poco irán saliendo cosas que evidentemente serán muy de andar por casa pero que pueden servir a quien las lea para aclarar los fundamentos y para incitarle la profundización personal. Puedo asegurar que en la investigación de libros y trabajos científicos le entrará un mareo del que no se sale ni tomando aspirinas.
De todas formas es casi imposible saber lo que pasó hace millones de años y mucho menos reproducirlo en un laboratorio. De hecho para cada tema particular hay un buen puñado de científicos investigando y cada día sale una cosa nueva. Los datos y las observaciones son los que son, casi siempre incuestionables. Pero saltar a la descripción de lo que pasó… todo queda en el campo de la parsimonia y la probabilidad de ocurrencia. Parece que, pudo ser… No obstante yo me quedo asombrado de la capacidad de imaginación que le ha echado el hombre para poder llegar a conclusiones tan serias y fiables a partir de lo poco que observamos. Sólo tienes que oír a alguno de los tres primeros espadas de Atapuerca (Arsuaga, Carbonell o Bermúdez de Castro) lo que son capaces de intuir a la vista de un pedacito de hueso de hace 800.000 años. Y como ellos un millón de sabios más.
El Anomalocaris me cae simpático. Quizás por los magníficos dibujos que se han hecho de lo que pudo ser. Bien calvo y con ojos de huevo que era. Por desgracia aún no hemos encontrado restos fósiles o indicadores biológicos de los hombres lobo acuáticos que nos permita enhebrar una historia coherente que correlacione a ambas especies. Pero podemos imaginar… Con el Anomalocaris hay gente que dice que era un terrible depredador del Cámbrico y hay otros que dicen que comían minucias planctónicas y detritus de los fondos marinos y pacían en los tapetes bacterianos ¡¡¡Chi lo sa!!!
Te dejo que me he enrollado un montón. Hasta la próxima.
jreguart : encontré esto para compartir , lo encontré muy novedoso , y lo ubiqué creo que cerca del periodo que le corresponde en la escala geológica http://www.elmundo.es/ciencia/2015/12/22/56792ee046163f5f558b459e.html
Hola Kambrico,
muchas gracias por la información. Ya la había visto e incluso me ha dado tiempo de incorporarla a la entrada correspondiente. Creo que fue en la que trata de la evolución inicial de los diferentes filos, al hablar del “camino de los artrópodos”.
Entre todos vamos construyendo. De nuevo muchas gracias.
Escribe un comentario